More than two alleles
The colors in hair, skin, and eyes are caused by
melanin. Microscopic granules of melanin
are deposited in the hair shafts. The
shape, size, and arrangement of these granules affect coat color. There are two different kinds of melanin:
eumelanin and phaeomelanin. Eumelanin
granules are thought to be spherical in shape and absorb almost all light,
giving black pigmentation. Phaeomelanin
granules are thought to be elongated like footballs, and to reflect light in
the red-orange-yellow range.
Black
The black gene has
three alleles that control the density of eumelanin granules in the hair
shaft. The black allele, B, is
dominant and produces a black (actually super-dark brown) coat. This black color is evident in all-black
cats, the black stripes of a tabby cat, and the dark ear-tips, feet and tails
(the points) of seal point Siamese cats.
The dark-brown allele, b, reduces black to a dark brown,
chocolate color. This allele is less
common and occurs almost exclusively in pedigreed cats such as Siamese and
Burmese. It is believed that the b
allele migrated into Europe from Asia
via importation of Siamese cats during the 1800’s. The light-brown allele, bl,
further reduces melanin density to produce a medium brown coat, called
cinnamon. It is not too likely that you
will find cats homozygous for these recessive alleles in Berkeley’s
general cat population. The dominance
heirarchy of these alleles is B > b > bl (Figure 2).
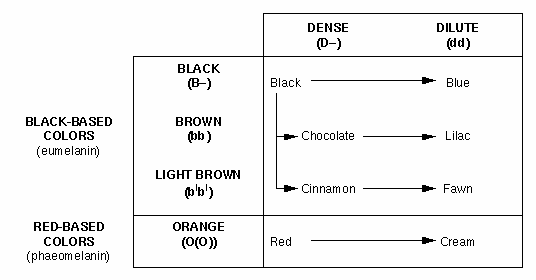
Dilute
The dilute locus
has two alleles that affect the distribution of pigment granules in the
fur. The dominant dense allele, D, produces
dense color, whereas the recessive dilute
allele (d) causes clumpng of pigment
granules in the hair shaft, leaving large areas between the clumps containing
no granules. These open areas cause
color dilution (Figure 2).
Figure 2. Effect of black
(eumelanin), orange (phaeomelanin),
and dilute genes on cat coat colors.
Codominance
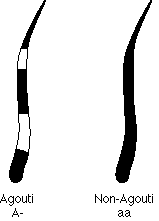
Dominance is a continuous phenomenon, which runs the gamut
from completely dominant (e.g. the shorthaired
allele, which completely masks the recessive longhaired allele) to codominant,
where the hetrozyogote resembles neither homozygote.
Albino
Two alleles at the albino locus, pointed, cs,
and sepia, cb, provide an example of codominance. Both alleles are recessive to the full-color allele, C, but are codominant with each other. The homozygous genotype cscs reduces pigment expression across most
of the animal. Reducing pigment
production in the eyes produces bright blue eyes, while reducing pigment
density in the hair shaft reduces the basic coat color from black/brown to a
light beige with dark brown “points” in the classic Siamese pattern. The homozygous cb cb genotype has a similar but smaller
reduction in pigment production, reducing a black coat to very dark brown
(similar to the color of the “points” on the Siamese) and yielding green or
green-gold eyes. These cats are called
“Burmese”. The heterozygote possesses
one of each of the alleles, cb cs,
and yields a combination phenotype called “Tonkinese”: a Siamese-patterned coat
with darker base body color and turquoise (aquamarine) eyes. All three alleles (C, cs, and cb) are dominant to the very
rare albino alleles, c and ca, which when homozygous
produce white cats with either lightly pigmented (caca) pale blue eyes or unpigmented (cc) pink eyes. The dominance hierarchy at the albino locus is: C > cb = cs > ca > c.
Question 3: Why do
population geneticists prefer to work with codominant genes?
Genotype by Environment Interaction
Most cat coat color traits show little influence from the
environment. However, there is one
fascinating exception to this rule. The Siamese allele, cs, at the albino
gene causes temperature-sensitive pigment expression. The allele produces a temperature-sensitive
tyrosinase gene that is inactive at the cat’s core body temperature, leaving a
light brown background. However, at the
tips of the extremities, which are much cooler, the enzyme is active and
produces normal amounts of pigment, creating the characteristic dark “points”
of the Siamese. Indeed, “indoor” Siamese
cats living in warm homes tend to be lighter than their “outdoor” compatriots,
which can become quite dark during cold winters. The cb
allele is also temperature sensitive, but less so than the cs allele, and therefore
produces a darker coat.
Epistasis: Gene-Gene Interaction
Orange
The orange gene
has two alleles: non- orange and orange.
The non-orange allele, o, is recessive and allows full
expression of the black locus. The dominant orange allele, O,
however, influences expression of the black
and agouti loci because it
substitutes the production of phaeomelanin for eumelanin. It masks the effect of the black gene by converting a black or
brown coat to orange. The ability of one
gene to mask the effect of another gene is called epistasis. Further, all
orange cats are tabbies because the orange
allele is epistatic to the non-agouti (solid coat) phenotype normally produced
by aa at the agouti locus. This masking
occurs because the orange band of phaeomelanin granules in the hair shaft is
not visible against the yellowish background of the hair without melanin
granules.
Sex-linked Traits
The other interesting characteristic of the orange gene is that it is carried on the
X chromosome, which makes it sex-linked. In male cats, this locus can normally produce
only two phenotypes, black or orange, whereas in females it can produce three
phenotypes: black, orange, and tortoiseshell.
This is because males are normally XY (heterogametic), and therefore have only one X-chromosome. Thus, if a male carries the orange allele at all, he will be orange
(OY).
Females are XX, meaning they have two X-chromosomes (homogametic). If both chromosomes carry the orange allele, then the cat will be
orange. However, if she is heterozygous
(Oo), her coat will be a patchwork of
orange and black patches, called tortoiseshell.
This pattern reveals an interesting phenomenon. To ensure that the amount of gene product in
female cells is equal to that found in male cells, early in female embryonic
development, one X-chromosome is inactivated in each cell. Thus, in a female that is heterozygous at the
orange locus, some cells produce
phaeomelanin (the active X-chromosome contains the O allele) and others do not (the active X-chromosome contains the o allele). Which X-chromosome is inactivated is an
entirely random decision. Further, this
decision is passed on to all cells in any cell line, making the female a mosaic
of cell patches with one or the other X-chromosomes actively producing its gene
products.
Question 4:
Develop a table describing the possible genotypes and their phenotypes that
could result from a cross between a tortoiseshell and a black cat. Assume both cats are homozygous BB.
Complex Traits
Complex traits are controlled by more than one gene. Of course, cat color is controlled by
multiple genes. However, even within
this complex trait, categories can be distinguished which allow us to ascertain
genotype at several loci. Some component
traits are themselves complex, probably because they are determined in part by
a locus with major effect, but also in part by loci with small effects. These loci with small effects are often
called minor loci or modifier genes.
Tabby
The tabby gene
causes banded (ticked) hairs to alternate with stripes, blotches, or spots of
solidly colored hairs, creating the striping pattern in cat coats. There are two common striping patterns
(Figure 3): mackerel (parallel stripes) or classic (characterized by thick
stripes or whorls that create a blotched or bulls-eye pattern). The parallel
stripe is produced by the dominant tabby
allele, T. It is probably the ancestral striping
pattern, which is seen in the African wild cat (Felis libyca) and the European wild cat (Felis silvestris). The
recessive blotched allele, tb, produces the classic
pattern, and probably arose much later by mutation. A third allele in the series, Abyssinian (Ta) produces the Abyssinian phenotype that has faint
striping only on the face or tail and has a dark stripe down the center of the
back. The hierarchy of dominance for the
three alleles is Ta > T > tb. However, Ta allele shows some degree
of incomplete dominance since either TaT
or Tatb
heterozygotes may show faint striping on the legs and tail.
Question 5: What effect might a striped (vs. solid) coat
have on the fitness of a wild cat?
Figure 3. Striping patterns determined by the tabby gene.

T‑ tbtb Ta‑
All cats have one of the above tabby patterns - even solid
black cats. However, because the agouti
gene is epistatic to the tabby gene,
striping cannot be seen in non-agouti cats.
This is because the recessive aa
agouti genotype blocks the production of banded hairs, thereby masking the
expression of the tabby phenotype. It is
nevertheless sometimes possible to see “ghost” striping on black cats,
especially kittens, when viewed in the proper light. It is also important to note that because the
O allele at the orange locus is epistatic to the agouti gene, tabby striping is not masked on solid orange cats, or
on orange patches on tortoiseshell cats.
That is, O is epistatic to the
epistatis of aa on the tabby gene.
The genetic mechanism behind a fourth tabby phenotype, the
spotted tabby, is not completely understood.
There are probably two different mechanisms. The first appears to be a modification of the
mackerel tabby, in which the narrow bars are broken up into small dark
patches. This effect is though to be the
result of several modifier genes acting on the major tabby gene. There is another rarer spotted phenotype
called the Ocicat (because of its resemblance to the wild ocelot) which has
larger, more distinct round spots. This
phenotype may be due to a separate major gene.
However, because it is so rare, for the purposes of this project we will
consider any spotted tabby to have the mackerel tabby phenotype, with
modifications.
Pleiotropy
When a gene affects more than one trait, it is termed pleiotropic. Most coat color genes have pleiotropic
effects on eye color. However, two genes
also have an interesting pleiotropic effect on hearing ability.
Dominant
White
The dominant white
allele, W, overrides all other genes
for pigmentation, and produces a white coat and blue eyes. Thus, the dominant white allele is epistatic to all other coat color genes. Of course, the other genes for color and
pattern are present, but their effects are completely hidden because the
dominant white mutation blocks
production of specialized melanin-producing cells called melanocytes.
Interestingly, the cochlea of the ear contains a band of
melanocytes that regulate ion balance.
Normally, hearing occurs via electrical signals stimulated by vibration
of the hair cells in the cochlea.
Transmission of these signals to the brain requires constant regulation
of ion balance. When ion balance is not
maintained, signal transmission to the brain degenerates within a few days
after birth, producing irreversible deafness.
Thus, the dominant white locus exhibits plieotropy,
affecting both coat color and hearing.
Piebald
Spotting
White spotted, or piebald, cats are very common. Spotting may occur with any coat color, and
is mainly due to the effect of one gene with two co-dominant alleles. The
spotted allele, S, creates
responsible for white spots, while the s
allele is not. Hence, the homozygote, ss, has no white spots. The heterozygote, Ss, has restricted areas of white spotting; usually the feet, nose,
chest, and belly. Finally, the SS homozygote has white regions covering
more than half the body. In the latter
case, people usually consider the dark patches to be spots. However, in reality, it is the larger white
area that is the spot, and in fact, a “spotted” (SS) cat may even be completely white!
The degree of spotting varies tremendously, but spotting
patterns usually follow a regular progression.
Cats with the least spotting have small spots on the breast and
belly. Increased spotting seems to
progress to cover the entire belly, the neck, chin and front feet. Finally, cats with the most spotting have
spots up the sides, over the back and onto the head. The tail seems to be the last area to have
white spots. Since cats vary
continuously in the extent of spotting, and the pattern of spotting is not
completely random, it is unlikely that only one gene determines the degree of
spotting. There is evidence of at least
one other gene that has a weak spotting effect.
It can cause a very small white spot on the throat, breast, or on the
belly near the hind legs. In fact,
several genes probably modify the action of the major spotting gene to produce
the continuum of spotting patterns seen in cat populations. Since the action of these weak modifier genes
is not well understood, we will score only the major spotting gene in our cat
survey.
The spotted allele, S,
hampers the migration of melanocytes during embryonic development. White spots are areas lacking
melanocytes. Thus, within these patches
a spotted cat will exhibit the same pleiotropy as caused by the dominant-white gene. For example, if an eye is within a spot, it
will be blue. Thus, spotted cats may be
blue-eyed or odd-eyed. Further, if the
spot encompasses an ear, the cat will be deaf in that ear. Since a spot often covers the eye when it
covers the ear, an odd-eyed cat will frequently be deaf on the blue-eyed
side.
The spotting gene has in interesting effect on the
patterning in tortoiseshell (Oo)
cats. With the ss genotype, there is no white spotting and the orange and black
are intermingled, usually without large patches of either orange or black. With either Ss or SS genotypes, there
are white spots and the orange and black occur as distinct patches. These cats are called calico cats (or,
sometimes, tortoiseshell and white).
This pattern seems to be due to the effect of the spotting allele on
embryonic melanocyte migration.
Special note about white cats
White cats can be produced in three distinct ways. It can be dominant white (W‑), albino (cc), or spotted (S‑)
with one big white spot all over its body.
It is therefore important to observe white cats carefully and take notes
about their eye-color.
The dominant white
allele, W, and the homozygous spotted
genotype, SS, can both produce a translucent
all-white coat with either orange or medium blue eyes. However, the spotted white cat often has a
tuft of color somewhere on its body. If
the cat has even one colored hair on its body, it is spotted rather than
dominant white
The albino gene
controls the amount of pigment produced in the melanocytes. A cat homozygous for albino, cc, at the albino locus is a true albino, in that
its melanocytes cannot produce any pigment.
It has a translucent white coat and pink eyes. A caca
genotype at the albino locus produces
a white cat with pale blue (almost grey) eyes.
Both these alleles are very rare.
Deafness in white cats is associated with white spotting (S), and with dominant white (W), but not with albino white (cc or caca).
Question 6: If you mated two white cats, would you be
confident of always obtaining white progeny?
Explain.
Question 7: What factors determine allele frequency? Is a dominant allele always the most frequent
allele at a locus?
Question 8: Manx cats have no tail and relatively long
rear legs. What is the genotype of a
Manx cat? Do you expect the frequency of
Manx cats to increase over time? Why or
why not?
_____________________________________________________________________________
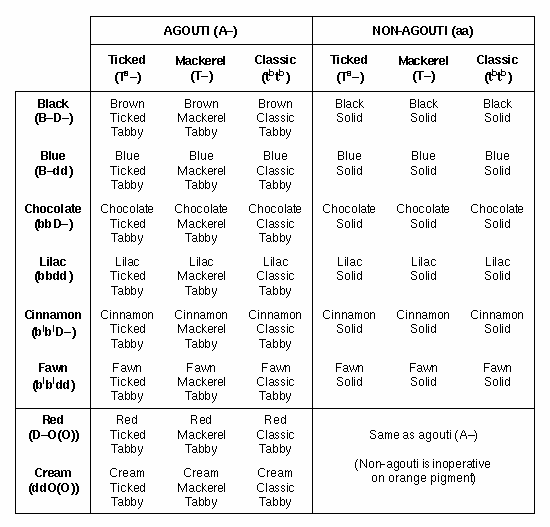
Table
2. Effects of agouti and tabby on the black, dilute and orange gene
effects. Note that here, an orange cat
is described as “red”, and an Abysinnian is called a “ticked tabby”. The agouti
and tabby genes also influence
expression of the albino gene, but their effects have not been tabulated
here.