Approach to the modern synthesis
Timeline of historical events in biology
http://www.biologie.uni-hamburg.de/b-online/e01/geschichte.htm
Timeline of historical events in genetics
http://las.alfred.edu/~emmonsc/Site/bio365/timeline.htm
Mendel’s dihybrid crosses
Mendel also did crosses where he kept track of two characters
Dihybrid (two-factor) crosses
Found that when he followed two traits, they were inherited independently
Crossed yellow/round x green/wrinkled
All progeny in F1 generation were yellow/round
Diagram:
GGWW x ggww = GgWw
Also crossed yellow/wrinkled x green/round
Again, all progeny in F1 were yellow/round
Diagram:
GGww x ggWW = GgWw
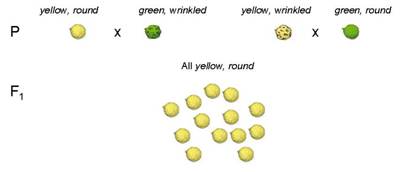
To produce F2 generation
Cross two individuals from the F1
Got this wild outcome

What creates this collection and ratio of phenotypes?
Mendel postulated that seed coat color and seed coat texture were inherited independently
So, used Product Rule to predict phenotype frequencies
Can do this in two ways
First method: Predicting independent phenotype frequencies
Summary
Use Product Rule and Punnett square for each monohybrid cross to predict independent phenotype frequencies
Then use Product Rule to predict joint phenotype frequencies
Steps
First, use monohybrid cross for coat color or predict coat color phenotypes
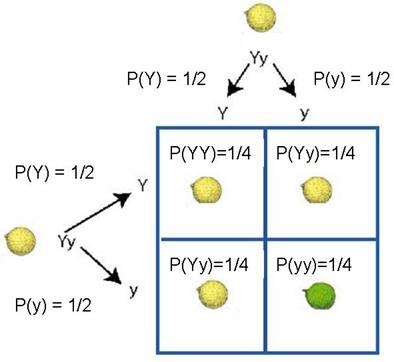
Then, use Summation Rule to obtain phenotype frequencies
P(Yellow) = ¼ + ¼ + ¼ = ¾
P(Green) = ¼
Next, use monohybrid cross for coat texture to predict coat texture phenotypes
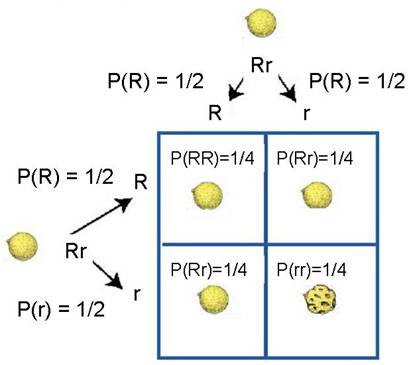
Then, use Summation Rule to obtain phenotype frequencies
P(Round) = ¼ + ¼ + ¼ = ¾
P(Wrinkled) = ¼
Then use Product Rule to predict joint phenotype frequencies
¾ yellow x ¾ round = 9/16 round yellow
¼ green x ¾ round = 3/16 round green
etc.
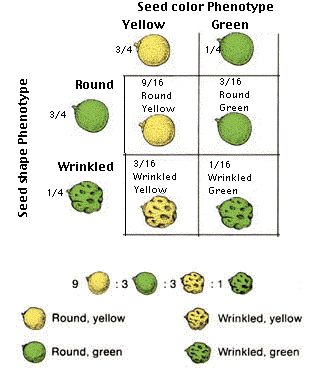
Here’s another way of illustrating this same process:
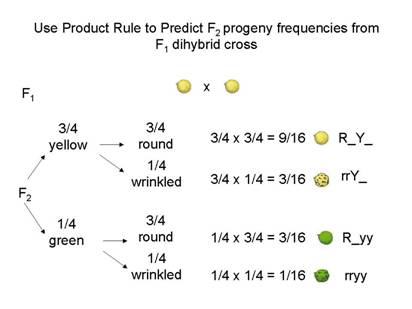
Second method: Predicting joint gamete frequencies
Summary
Use Product Rule to predict joint gamete frequencies
Use Product Rule to predict phenotype frequencies
Steps
Use Product Rule to predict joint gamete frequencies
Possible gametes from RrYy
P(RY gamete) = ½ R x ½ Y = ¼
P(Ry gamete) = ½ R x ½ y = ¼
P(rY gamete) = ½ r x ½ Y = ¼
P(ry gamete) = ½ r x ½ y = ¼
Use joint gamete frequencies to generate a large Punnet
Square
Punnett square uses Product Rule to predict genotype frequencies
e.g. RY fuses with RY to produce RRYY
Therefore P(RRYY) = P(RY) x P(RY) = ¼ x ¼ = 1/16
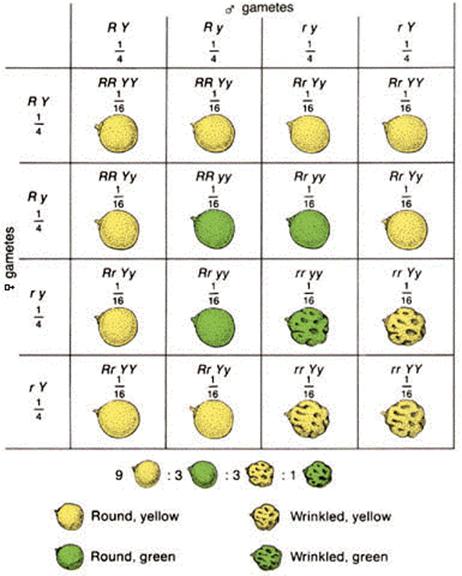
Then use Summation Rule to predict phenotype frequencies
P(Round, yellow) = P(R-Y-) = P(RRYY) + P(RrYY) + P(RRYy) + P(RrYy) = 1/16 + 1/16 + 1/16 + 1/16 + 1/16 + 1/16 + 1/16 + 1/16 +1/16 + 1/16 = 9/16
Mendel’s Fourth Postulate
All these methods depend on Mendel’s Fourth Postulate
Mendel observed that each trait was inherited independently
Followed Product Law
Independent assortment of heritable units
Later, we’ll discuss situations in which traits are not strictly independently inherited
But, this is a good predictive model
Any observed mathematical deviations from the model point to situations that need more investigations
As we will see shortly
To test your understanding, try some dihybrid cross problems:
http://www.biology.arizona.edu/mendelian_genetics/problem_sets/dihybrid_cross/dihybrid_cross.html
Chromosomal Theory of Inheritance
1902-1903 Walter Sutton and Theodor Boveri
Noted parallelism between chromosome behavior and Mendelism
Precise sorting
and recombination of chromosomes in formation of germ cells was striking
Independently
proposed that each egg or sperm cell contains only one member of each
chromosome pair
Mendel postulated that each individual has two copies of each heritable unit controlling a trait, one inherited from each parent
Likewise, Sutton and Boveri both observed that each individual has two copies of each type of chromosome
A gamete
receives only one member of each chromosomal pair
Which would
allow progeny to receive one heritable unit from each parent
1902 Walter S. Sutton (1877 - 1916)
While a graduate student in E. B. Wilson’s lab at Columbia
University
Observed homologous pairs of chromosomes in grasshopper cells
These grasshopper chromosomes have quite distinct shapes, so can follow individual chromosomal pairs
Found that during meiosis the chromosome pairs split, and each chromosome goes to its own cell
The segregation pattern of chromosomes during meiosis matched the segregation patterns of Mendel’s genes.
He suggested that Mendel's "factors" must be located on chromosomes
"…the association of paternal and maternal chromosomes in pairs and their subsequent separation during the reducing division as indicated above may constitute the physical basis of the Mendelian law of heredity" (Sutton, 1902).
Sutton W.S. 1902. On the morphology of the chromosome group in Bracystola magna. Biol. Bull. 4:24-39.
In 1903, he made an even stronger argument to connect Mendel’s laws of heredity and the behavior of chromosomes in his paper: The Chromosomes in Heredity.
Sutton reiterated earlier work of Theodor Boveri, who in the late 1880s and early 1890s observed that chromosome numbers are cut in half as egg cells mature, and concluded that sperm and egg nuclei have half sets of chromosomes.
1902 Boveri
Discovered that a particular combination of chromosomes, rather than their number, is essential for normal cel development
Abnormal fertilization of sea urchin eggs by two sperm cells causes unequal distribution of chromosomes in resulting daughter cells
For example, one copy of chromosome c is attached to poles 1 and 2, and one copy is attached to poles 2 and 3
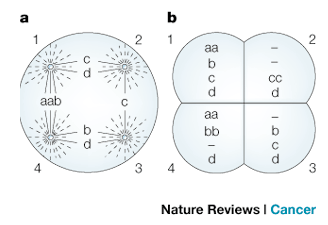
Thus, when chromosomes are segregated to the four poles at cell division, some daughter cells have too many copies of certain chromosomes and some have too few
For example, cell 2 has two copies of chromosome c and cell 4 has none.
Boveri found that these abnormal zygotes could not complete development
Today’s view of chromosomal inheritance
Single molecule of DNA in each sister chromatid
During meiosis and mitosis, packaged into four-armed structure
Each set of arms (a short and a long) is a chromatid
Mitosis
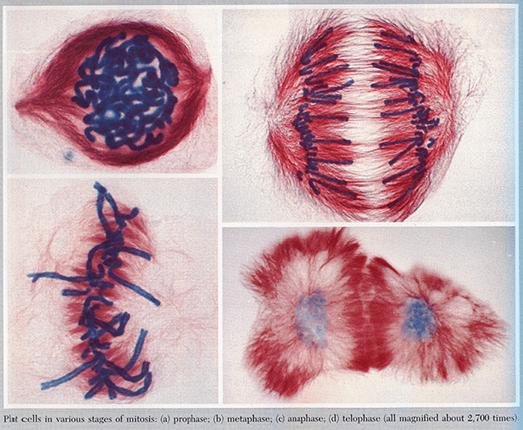
In mitosis, the pairs duplicate and one set goes to each progeny cell
Giving the daughter cells a complete complement of all the information in the parent cell
Mitotic division is used for reproduction by single-celled organisms
Hence, they are clones of one another
A lineage of bacteria are essentially genetically identical
Mitosis provides for cell proliferation during growth in a multicellular organism
Gametogenesis
However, in gametogenesis, the number of chromosomes must be reduced. Why?
To avoid accumulation of chromosomes during fertilization
Meiosis

Gemete formation in dihybrid cross
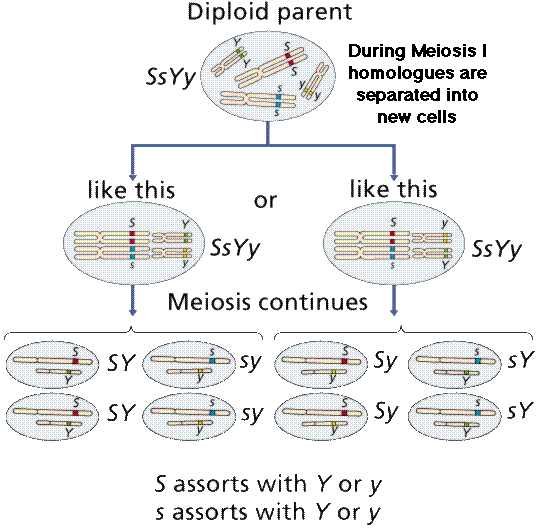
Important points
First meiotic prophase
Every 4-armed chromosome pairs up with its homolog
One homolog was from mother, one was from father
First meiotic anaphase
Homologs segregate to one side or other of cell
Non-homologous chromosomes assort independently
In some pairs, father’s copy goes to right
In some other pairs, mother’s copy goes to right
Second meiotic anaphase
Chromatids separate
Every gamete gets a complete set (one member of each pair)
So, at fertilization, every chromosome will have a homolog (back to two units)
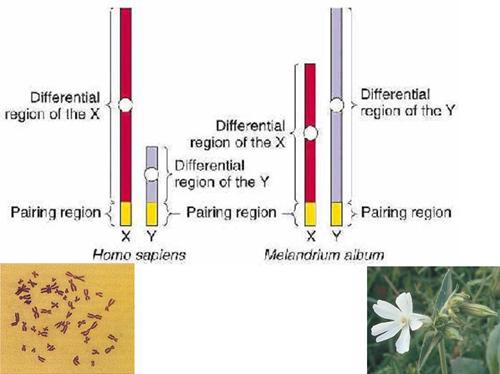
Chromosomal sex-determination
1905 Edmund Beecher Wilson
Studying insects, independently propose that separate X and Y chromosomes determine sex.
Stevens, studying the scarab Tenebrio molitor shows that a single Y chromosome determines maleness, and two copies of the X chromosome determine femaleness.
Her work on sex determination was published as a Carnegie Institute report in 1905. She died in 1912 of breast cancer.
Biographical info: http://www.dnaftb.org/dnaftb/concept_9/con9bio.html
In many organisms, sexes are chromosomally determined
Homogametic sex -- that sex containing two like sex chromosomes
In most animals species these are females (XX)
Butterflies and Birds, ZZ males
Heterogametic sex --- that sex containing two different sex chromosomes
In most animal species these are XY males
Butterflies and birds, ZW females
Grasshopers have XO males
Sex determination in mammals
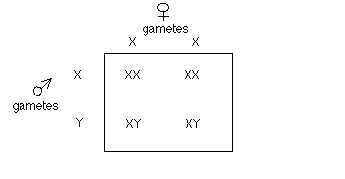
Patterns of sex linkage:
(a) Male, female progeny (sometimes) differ.
(b) Reciprocal crosses differ.
(c) For genes on the X chromosome, recessive alleles "appear" more often in males.
Punnett was a poultry geneticist
During the First World War, Punnett developed a technique of separating male and female chicks using sex-linked plumage colours.
In this way, the less useful male chicks could be separated from the more useful female chicks and destroyed.
This improved the efficiency of the poultry industry.
The first autosexing breed developed was the Cambar back in 1929 (Gold Campine X Barred Rock).
Barred plumage (alternating
black and white bands on feathers) is a dominant sex-linked trait in chickens.
Remember, in birds, males
are heterogametic
But, because barred is dominant, a male needs only
one copy of the barred allele for its plumage to be barred
Punnett
developed a breeding strategy using sex-linked plumage color to differentiate
between newly-hatched male and female chicks
In Punnett’s strategy, the female parent must carry the dominant allele for the sex-linked trait
In this case, all male progeny get a recessive non-barred allele from their father, but the dominant barred allele from their mother makes them barred
Female progeny get only the non-barred allele from their father, and so are non-barred
Pictures illustrate the Cream Legbar breed
Leghorn
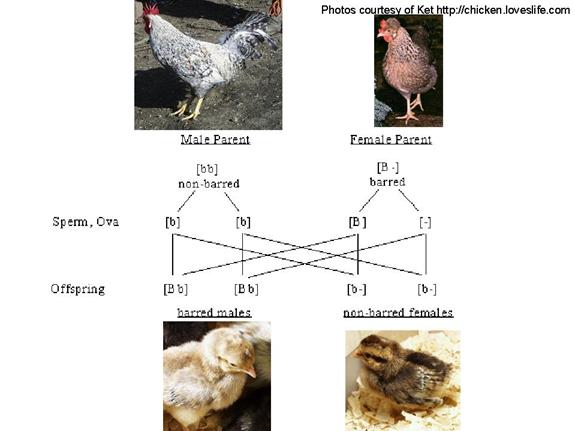
As an exercise:
Try doing a Punnet square for this cross
Then, do a Punnet square for the reciprocal cross (where mother is non-barred and father is barred)
Homogametic sex is mosaic
Dosage compensation – shut down one copy of long arm
Random from one cell lineage to another
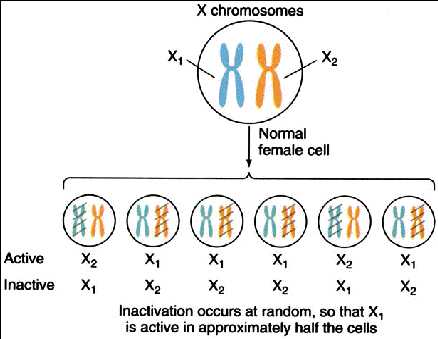
Creates patchiness – mosaic
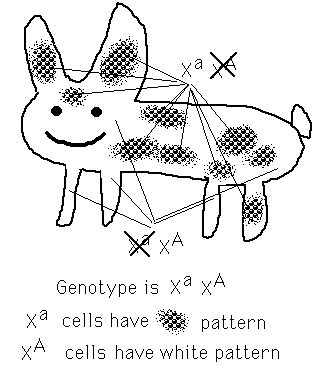
e.g. in cats, orange locus on sex chromosome
Females are heterogametic Oo
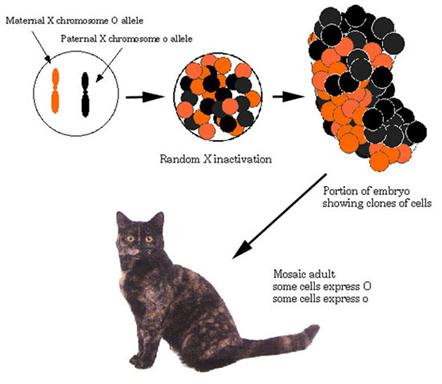
Leading to a tortoiseshell cat
Only way to get completely orange cat is for it to be male: O-
For more details, see handout on cat coat-color genetics (Rich make a link here)

Gene interaction (epistasis)
1905 William Bateson and Reginald Crundall Punnett (1875–1967)
Found various deviations from the normal dihybrid ratio (9:3:3:1), which they rightly attributed to gene interaction.
They analyzed the three comb types of chicken known to exist at that time:
Chicken Varieties |
Phenotype |
|
Rose Comb |
Brahmas |
Pea Comb |
Leghorns |
Single Comb |
Result
The F1 differed
from both parents
New, walnut
phenotype
Two new
phenotypes not seen in the parents appeared in the F2
Analysis
How can this
result be explained?
Work this out at
your desk
Talk to your neighbors
Clues:
The first clue
is the F2 ratio (9:3:3:1)
Same ratio as F1 from a sib crossing of dihybrid cross progeny
This observation suggests that two genes may control the phenotype of the comb
Make a dihybrid cross
A series of
experiments demonstrated that the genotypes controlling the various comb
phenotypes were as follows.
Phenotypes
|
Genotypes
|
Frequency
|
Walnut
|
R_P_
|
9/16
|
Rose
|
R_pp
|
3/16
|
Pea
|
rrP_
|
3/16
|
Single
|
rrpp
|
1/16
|
What test
crosses are needed to determine parental phenotypes?
The gene interactions and genotypes were determined by performing the appropriate testcrosses
Two possible genotypes for each parent
Rose: RRpp
Pea: rrPP
Or
Rose Rrpp
Pea: rrPp
It was later shown that the genotypes of the initial parents were:
Rose = RRpp
Pea = rrPP
Therefore,
genotypically the cross was:
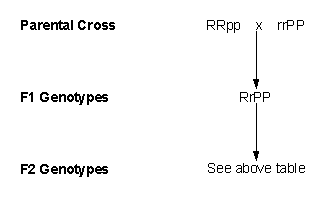
|
Walnut |
R_P_ |
9/16 |
|
Rose |
R_pp |
3/16 |
|
Pea |
rrP_ |
3/16 |
|
Single |
rrpp |
1/16 |